
-

����ͨ��
����ץס�����Ƽ�
���ӵ��}��
�ע��tRNA�о���PCRоƬ����nrStar™ Human tRNA Repertoire PCR Array
�����w�� �� �� С �� �r�g��2016��12��08�� ��Դ����������
�����]��
����Arraystar���°l�����Ј����ע��tRNA�о���PCRоƬ����nrStar™ Human tRNA Repertoire PCR Array��ԓ��оƬ��ͬ�r�z�y66��������tRNA��22�������wtRNA�����w��tRNA����������GtRNAdb��tRNAdb�е����з��ܴa�ӣ�����͑����ٵ��M��tRNA���_�V������
�D�\RNA (tRNA) �������w�ȷֲ���V�����������S���ķǾ��aСRNA���ӡ����y�����D�\�����ᣬ���c�����g�����B��mRNA�c�����|����Ҫ������������ֳ[1]���ֻ�[1, 2]�͵���[3]��һϵ������W�^�̶����S��tRNAˮƽ��׃������֮��tRNA���_�V�ĸ�׃Ҳ��Ӱ푼����l���^���е����\����_ʧ�{��tRNA���Դ��M�[���İl���Ͱ��Y�M��[2, 4-11]�����⣬�S��������������II������[12]����͢�D�Y[13]�Լ�HIV��Ⱦ[14]�����F��tRNA���_�c�ֲ��Ɂy��tRNA�о�����u�ɞ�����W�^�̺ͼ����о�����Ҫ�M�ɲ��֡�
Arraystar���°l�����Ј����ע��tRNA�о���PCRоƬ����nrStar™ Human tRNA Repertoire PCR Array��ԓ��оƬ��ͬ�r�z�y66��������tRNA��22�������wtRNA�����w��tRNA����������GtRNAdb��tRNAdb�е����з��ܴa�ӣ�����͑����ٵ��M��tRNA���_�V����������һ�����ܴa��ƫ���ԡ������gЧ�ʺ͜ʴ_�ԡ����������W�Լ�������Ⱦ�ļ������Ե��о��ṩ��Ҫ���������M��tRNA�о��ز����ٵĹ��ߡ�
tRNA���ڷN�����������l�]����ʮ���P�I��Ȼ�����@Щ��������Ǽ������������K���D䛵��M�У�����cDNA�ϳɽKֹ��A���e�䡣��ˣ�Arraystar���T�_�l��ᘌ�tRNA�ķ��D�ԇ����(rtStar™ tRNA-optimized First-Strand Synthesis Kit)��ԓԇ���в�����һ�N��Ч��RNAȥ����øAlkB���܉���Ч��ȥ��tRNA�ϵļ�������O������cDNA�ϳ��|�����c��ԇ���нM��ʹ�ã��о��ˆT�܉�@�ø����挍�ɿ���tRNA���_׃�������о������|�M��tRNA��ԴƬ�Σ�tRFs���ṩ��Ҫ��Ϣ��
���˱��C�������Ŷȣ�оƬ߀������3������СRNA����ȅ����Լ�3���|�،���RNA Spike-in��PCR��Ԍ��գ�PPC���ͻ���MDNA���գ�GDC��,�քe���ڱO�ycDNA�ϳ��|����PCRЧ�ʺͻ���MDNA��Ⱦ��
�ں��m�о��У��õ�tRNA[15] ���^���_tRNA [2, 16, 17]�����Mһ����ʾ�ض�tRNA�ڼ�����������е����á��������ڷǾ��aRNA�о������P���gҲ�m����tRNA�ĺ��m�о���
�aƷ�б�
|
�aƷ���Q |
Ҏ�� |
���� |
|
nrStar™ Human tRNA Repertoire PCR Array |
384-well (4*96) plate |
����66��������tRNA��22�������wtRNA |
оƬ���c
• ����GtRNAdb��tRNAdb�����������еļ������c�����w���ܴa��
• ���S��ȥ����̎��ʹ�Ùz�y�Y�������挍�ɿ�
• ����������ڶ�N�����ͽM����ͨ�^��C
• ��������384�װ壬��С�r�ȱ�ɵõ��Y��
��������
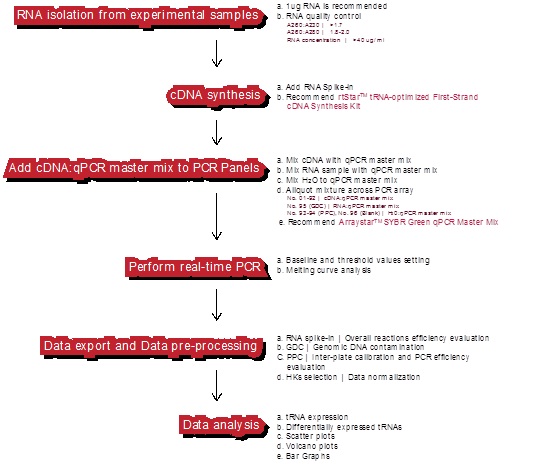
�m�Ì��r�����ɹ�PCR�x��
ABI ViiA™ 7��ABI 7500 & ABI 7500 FAST��ABI 7900HT��ABI QuantStudio™ 6 Flex Real-Time PCR system��ABI QuantStudio™ 7 Flex Real-Time PCR system��ABI QuantStudio™ 12K Flex Real-Time PCR System��Bio-Rad CFX384��Bio-Rad iCycler & iQ Real-Time PCR Systems��Eppendorf Realplex��QIAGEN Rotor Gene Q100��Roche LightCycler 480��Stratagene Mx3000��Roche LightCycler 480
������
tRNA���ܴa����Ϣ(GtRNAdb��tRNAdb������)��
|
������tRNA (66): Ala-AGC, Ala-CGC, Ala-GGC, Ala-TGC, Arg-ACG, Arg-CCG, Arg-CCT, Arg-TCG, Arg-TCT, Asn-ATT, Asn-GTT, Asp-ATC, Asp-GTC, Cys-GCA, Gln-CTG-1, Gln-CTG-2, Gln-TTG-1, Gln-TTG-2, Glu-CTC, Glu-TTC, Gly-CCC-1, Gly-CCC-2, Gly-GCC, Gly-TCC, His-GTG, Ile-AAT, Ile-TAT, Ini-CAT, Leu-AAG, Leu-CAA, Leu-CAG, Leu-TAA, Leu-TAG, Lys-CTT-1, Lys-CTT-2, Lys-TTT, Met-CAT, Phe-GAA, Pro-AGG, Pro-CGG, Pro-GGG, Pro-TGG, Sec-TCA, Ser-ACT, Ser-AGA, Ser-CGA, Ser-GCT, Ser-GGA, Ser-TGA, Sup-CTA, Sup-TTA, Thr-AGT-1, Thr-AGT-2, Thr-CGT, Thr-TGT-1, Thr-TGT-2, Trp-CCA, Tyr-ATA, Tyr-GAT, Tyr-GTA, Val-AAC, Val-CAC-1, Val-CAC-2, Val-CAC-3, Val-TAC-1, Val-TAC-2 |
|
�����wtRNA (22): mt-Ala-TGC, mt-Arg-TCG, mt-Asn-GTT, mt-Asp-GTC, mt-Cys-GCA, mt-Gln-TTG, mt-Glu-TTC, mt-Gly-TCC, mt-His-GTG, mt-Ile-GAT, mt-Leu-TAA, mt-Leu-TAG, mt-Lys-TTT, mt-Met-CAT, mt-Phe-GAA, mt-Pro-TGG, mt-Ser-GCT, mt-Ser-TGA, mt-Thr-TGT, mt-Trp-TCA, mt-Tyr-GTA, mt-Val-TAC |
������ȡnrStar Human tRNA Repertoire PCR Array��Ԕ���Y��
�����īI
[1] Gingold H, Tehler D, Christoffersen NR, Nielsen MM, Asmar F, Kooistra SM, et al. A dual program for translation regulation in cellular proliferation and differentiation. Cell 2014;158:1281-92.
[2] Pavon-Eternod M, Gomes S, Rosner MR, Pan T. Overexpression of initiator methionine tRNA leads to global reprogramming of tRNA expression and increased proliferation in human epithelial cells. Rna 2013;19:461-6.
[3] Mei Y, Stonestrom A, Hou YM, Yang X. Apoptotic regulation and tRNA. Protein & cell 2010;1:795-801.
[4] Berns A. A tRNA with oncogenic capacity. Cell 2008;133:29-30.
[5] Waldman YY, Tuller T, Sharan R, Ruppin E. TP53 cancerous mutations exhibit selection for translation efficiency. Cancer research 2009;69:8807-13.
[6] Kushner JP, Boll D, Quagliana J, Dickman S. Elevated methionine-tRNA synthetase activity in human colon cancer. Proceedings of the Society for Experimental Biology and Medicine Society for Experimental Biology and Medicine 1976;153:273-6.
[7] Marshall L, Kenneth NS, White RJ. Elevated tRNA(iMet) synthesis can drive cell proliferation and oncogenic transformation. Cell 2008;133:78-89.
[8] Pavon-Eternod M, Gomes S, Geslain R, Dai Q, Rosner MR, Pan T. tRNA over-expression in breast cancer and functional consequences. Nucleic acids research 2009;37:7268-80.
[9] Zhou Y, Goodenbour JM, Godley LA, Wickrema A, Pan T. High levels of tRNA abundance and alteration of tRNA charging by bortezomib in multiple myeloma. Biochemical and biophysical research communications 2009;385:160-4.
[10] Begley U, Sosa MS, Avivar-Valderas A, Patil A, Endres L, Estrada Y, et al. A human tRNA methyltransferase 9-like protein prevents tumour growth by regulating LIN9 and HIF1-alpha. EMBO molecular medicine 2013;5:366-83.
[11] Goodarzi H, Nguyen HC, Zhang S, Dill BD, Molina H, Tavazoie SF. Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression. Cell 2016;165:1416-27.
[12] Krokowski D, Han J, Saikia M, Majumder M, Yuan CL, Guan BJ, et al. A self-defeating anabolic program leads to beta-cell apoptosis in endoplasmic reticulum stress-induced diabetes via regulation of amino acid flux. The Journal of biological chemistry 2013;288:17202-13.
[13] Girstmair H, Saffert P, Rode S, Czech A, Holland G, Bannert N, et al. Depletion of cognate charged transfer RNA causes translational frameshifting within the expanded CAG stretch in huntingtin. Cell reports 2013;3:148-59.
[14] van Weringh A, Ragonnet-Cronin M, Pranckeviciene E, Pavon-Eternod M, Kleiman L, Xia X. HIV-1 modulates the tRNA pool to improve translation efficiency. Molecular biology and evolution 2011;28:1827-34.
[15] Fu G, Xu T, Shi Y, Wei N, Yang XL. tRNA-controlled nuclear import of a human tRNA synthetase. The Journal of biological chemistry 2012;287:9330-4.
[16] Gong M, Gong F, Yanofsky C. Overexpression of tnaC of Escherichia coli inhibits growth by depleting tRNA2Pro availability. Journal of bacteriology 2006;188:1892-8.
[17] Yona AH, Bloom-Ackermann Z, Frumkin I, Hanson-Smith V, Charpak-Amikam Y, Feng Q, et al. tRNA genes rapidly change in evolution to meet novel translational demands. eLife 2013;2:e01339.




